Генетический код
Большая Советская Энциклопедия. Статьи для написания рефератов, курсовых работ, научные статьи, биографии, очерки, аннотации, описания.
|
|
| ГЕБ |
| ГЕВ |
| ГЕГ |
| ГЕД |
| ГЕЕ |
| ГЕЗ |
| ГЕЙ |
| ГЕК |
| ГЕЛ |
| ГЕМ |
| ГЕН |
| ГЕО |
| ГЕП |
| ГЕР |
| ГЕС |
| ГЕТ |
| ГЕФ |
| ГЕЧ |
| ГЕШ |
| ГЕЯ |
Генетический код, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах — дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) — встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания, Пиримидиновые основания). В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых кислот обнаружено около 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных сахаров. Т. к. количество кодирующих знаков Генетический код к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. количество нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42=16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Генетический код Гамов предложил (1954) модель триплетного Генетический код к., т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, наз. кодоном. Число возможных триплетов равно 43=64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (т. н. вырожденность кода). Было предложено много различных моделей Генетический код к., из которых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены следующие основные закономерности, касающиеся Генетический код к.: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Генетический код к.); 2) считывание Генетический код к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Генетический код к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Генетический код к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за некоторыми исключениями). Универсальность Генетический код к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (например, кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (например, проростков гороха), то в такой системе, будет идти белковый синтез. Благодаря работам американских генетиков М. Ниренберга, С. Очоа, Х. Корана известен не только состав, но и порядок нуклеотидов во всех кодонах (см. табл., построенную по данным опытов с кишечной палочкой).
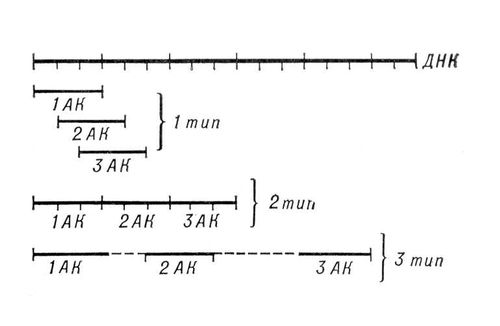
Модели генетического кода: 1-й тип — перекрывающийся код без запятых; 2-й тип — неперекрывающийся код без запятых; 3-й тип — код с «промежутками», т. е. код с запятыми.
Из 64 кодонов у бактерий и фагов 3 кодона — УАА, УАГ и УГА — не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их назыают терминирующими кодонами. Существуют также 3 сигнала о начале синтеза — это т. н. инициирующие кодоны — АУГ, ГУГ и УУГ, — которые, будучи включенными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида.
Реализация Генетический код к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит название транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК «переписывается» в нуклеотидную последовательность РНК. Второй этап — трансляция — протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке: этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки, раздел Биосинтез).
Полный «словарь» генетического кода для аминокислот
|
Первая «буква» | Вторая буква |
Третья «буква» | |||||||
| У |
Ц |
А |
Г | ||||||
У | УУУ | Фенилаланин | УЦУ | Серин | УАУ | Тирозин | УГУ | Цистин | У |
| УУЦ | УЦЦ | УАЦ | УГЦ | Ц | ||||
| УУА | Лейцин | УЦА | УАА | Конец синтеза | УГА | Конец синтеза | А | |
| УУГ* | УЦГ | УАГ | УГГ | Триптофан | Г | |||
Ц | ЦУУ | Лейцин | ЦЦУ | Пролин | ЦАУ | Гистидин | ЦГУ | Аргинин | У |
| ЦУЦ | ЦЦЦ | ЦАЦ | ЦГЦ | Ц | ||||
| ЦУА | ЦЦА | ЦАА | Глутамин | ЦГА | А | |||
| ЦУГ | ЦЦГ | ЦАГ | ЦГГ | Г | ||||
А | АУУ | Изолейцин | АЦУ | Треонин | ААУ | Аспарагин | АГУ | Серин | У |
| АУЦ | АЦЦ | ААЦ | АГЦ | Ц | ||||
| АУА | АЦА | ААА | Лизин | АГА | Аргинин | А | ||
| АУГ* | Метионин | АЦГ | ААГ | АГГ | Г | |||
Г | ГУУ | Валин | ГЦУ | Аланин | ГАУ | Аспарагиновая кислота | ГГУ | Глицин | У |
| ГУЦ | ГЦЦ | ГАЦ | ГГЦ | Ц | ||||
| ГУА | ГЦА | ГАА | Глутаминовая кислота | ГГА | А | |||
| ГУГ* | ГЦГ | ГАГ | ГГГ | Г | ||||
* В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов.
Лит.: Общая природа генетического кода для белков, в сборнике: Молекулярная генетика, пер. с англ., М., 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ., М., 1964, с. 9—23; Ниренберг М., Генетический код (II), там же, с. 24—41; Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер. с англ., М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М. — Л., 1966; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П.. Общая генетика, М., 1970.
Н. П. Дубинин, В. Н. Сойфер.
|
Так же Вы можете узнать о... Таналык, река в Башкирской АССР, низовье — в Оренбургской области РСФСР, правый приток р. Завадовский Михаил Михайлович [17(29).7.1891, с. Синисало Гельмер-Райнер Нестерович (р. 14.6. Гуковский Григорий Александрович [18.4(1.5). Редан (воен.) Редан (франц. redan — уступ), открытое полевое фортификационное сооружение из двух фасов под углом 60—120°, выступающим в сторону противника (рис. Времяимпульсный датчик, устройство, преобразующее измеряемую (контролируемую) величину в импульс, длительность которого (интервал времени) пропорциональна времени действия входного сигнала. Платежи из прибыли в бюджет, один из важнейших источников формирования доходов государственного бюджета при социализме. Бордюр (франц. bordure, от bord — край), кромка, кайма, полоска, кривые или прямые полосы, обрамляющие что-либо (рисунок, текст и пр. Обучающаяся автоматическая система, обучаемая машина, самоприспосабливающаяся система, алгоритм управления которой изменяется в соответствии с оценкой результатов управления так, что с течением времени она улучшает свои характеристики и качество функционирования (см. Арвида (Arvida), город на юго-востоке Канады, на р. Мистакокариды (Mystacocarida), отряд (или подкласс) ракообразных. Эрг-Игиди, песчаная пустыня в Западной Сахаре (Алжир, Мавритания), между плато Дра на северо-западе и плато Эль-Эглаб на юго-востоке. Лис, Лейе (франц. Lys, флам. Leie), река во Франции и Бельгии. Центральнолесной заповедник, заповедник в юго-западной части Калининской области, на водоразделе Волги и Западной Двины, в 30 км к С. Концентратор акустический, устройство для увеличения интенсивности звука. Уньоро, Буньоро, государство в Восточной Африке, в районе Межозерья (на вост. Калифорнийский университет (California University), один из крупнейших вузов США, открыт в 1868 как официальное учебное заведение штата Калифорния на базе Калифорнийского колледжа, основан в 1855 в Окленде близ Сан-Франциско. Сутора, рыжая сутора (Suthora webbiana), птица семейства толстоклювых синиц отряда воробьиных. Ерухан (псевдоним; настоящие имя и фамилия Ерванд Срмакешханлян) (1870—1915), армянский писатель. |
|