Кора больших полушарий головного мозга
Большая Советская Энциклопедия. Статьи для написания рефератов, курсовых работ, научные статьи, биографии, очерки, аннотации, описания.
|
|
| КОА |
| КОБ |
| КОВ |
| КОГ |
| КОД |
| КОЖ |
| КОЗ |
| КОИ |
| КОЙ |
| КОК |
| КОЛ |
| КОМ |
| КОН |
| КОО |
| КОП |
| КОР |
| КОС |
| КОТ |
| КОУ |
| КОФ |
| КОХ |
| КОЦ |
| КОЧ |
| КОШ |
| КОЩ |
| КОЭ |
| КОЮ |
| КОЯ |
Кора больших полушарий головного мозга, слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга, развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности, хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см2.
Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними (рис. 1). Пространство между телами и отростками нервных клеток коры заполнено нейроглией и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон (рис. 2). Звездчатые клетки отличаются слабым развитием дендритов и мощным развитием аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением (см. Клетка). Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику (см. рис. 1 и 3).
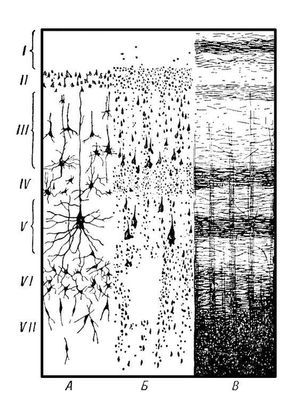
Рис. 1. Схема строения коры головного мозга человека: I — зональный слой, II — наружный зернистый слой, III — пирамидный слой, IV — внутренний зернистый слой, V — ганглионарный слой, VI — слой треугольных клеток, VII — слой веретеновидных клеток. А — нейронное строение, Б — цитоархитектоника, В — волоконная структура.
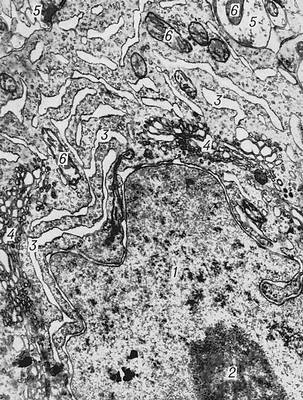
Рис. 2. Электронномикроскопическое строение пирамидной клетки коры головного мозга белой крысы: 1 — ядро; 2 — ядрышко; 3 — канальцы эндоплазматической сети; 4 — комплекс Гольджи; 5 — синапсы; 6 — митохондрии.
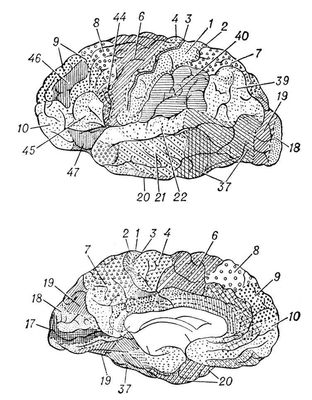
Рис. 3. Карта цитоархитектонических полей коры головного мозга человека: А — наружная поверхность полушария, Б — внутренняя поверхность полушария. Номерами и различной штриховкой обозначены цитоархитектонические поля коры.
Наиболее крупные подразделения территории коры — древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность новой коры у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%.
Если представить себе кору мозга в виде единого покрова (плаща), одевающего поверхность полушарий, то основная центральная часть его составит новая кора, в то время как древняя, старая и межуточная займут место на периферии, т. е. по краям этого плаща. Древняя кора у человека и высших млекопитающих состоит из одного клеточного слоя, нечетко отделённого от нижележащих подкорковых ядер; старая кора полностью отделена от последних и представлена 2—3 слоями; новая кора состоит, как правило, из 6—7 слоев клеток; межуточные формации — переходные структуры между полями старой и новой коры, а также древней и новой коры — из 4—5 слоев клеток. Неокортекс подразделяется на следующие области: прецентральную, постцентральную, височную, нижнетеменную, верхнетеменную, височно-теменно-затылочную, затылочную, островковую и лимбическую. В свою очередь, области подразделяются на подобласти и поля. Основной тип прямых и обратных связей новой коры — вертикальные пучки волокон, приносящие информацию из подкорковых структур к коре и посылающие её от коры в эти же подкорковые образования. Наряду с вертикальными связями имеются внутрикортикальные — горизонтальные — пучки ассоциативных волокон, проходящие на различных уровнях коры и в белом веществе под корой. Горизонтальные пучки наиболее характерны для I и III слоев коры, а в некоторых полях для V слоя. Горизонтальные пучки обеспечивают обмен информацией как между полями, расположенными на соседних извилинах, так и между отдалёнными участками коры (например, лобной и затылочной).
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений — нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры — комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора одна из колонок реагирует на прикосновение к коже, то другая — на движение конечности в суставе. В зрительном анализаторе функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя — в вертикальной и т. п.
Второй комплекс клеток новой коры — слой — ориентирован в горизонтальной плоскости. Полагают, что мелкоклеточные слои II и IV состоят в основном из воспринимающих элементов и являются «входами» в кору. Крупноклеточный слой V — выход из коры в подкорку, а среднеклеточный слой III — ассоциативный, связывающий между собой различные корковые зоны (см. рис. 1).
Локализация функций в коре характеризуется динамичностью в силу того, что, с одной стороны, имеются строго локализованные и пространственно отграниченные зоны коры, связанные с восприятием информации от определенного органа чувств, а с другой — кора является единым аппаратом, в котором отдельные структуры тесно связаны и в случае необходимости могут взаимозаменяться (т. н. пластичность корковых функций). Кроме того, в каждый данный момент корковые структуры (нейроны, поля, области) могут образовывать согласованно действующие комплексы, состав которых изменяется в зависимости от специфических и неспецифических стимулов, определяющих распределение торможения и возбуждения в коре. Наконец, существует тесная взаимозависимость между функциональным состоянием корковых зон и деятельностью подкорковых структур. Территории коры резко различаются по своим функциям. Большая часть древней коры входит в систему обонятельного анализатора. Старая и межуточная кора, будучи тесно связанными с древней корой как системами связей, так и эволюционно, не имеют прямого отношения к обонянию. Они входят в состав системы, ведающей регуляцией вегетативных реакций и эмоциональных состояний организма (см. Ретикулярная формация, Лимбическая система). Новая кора — совокупность конечных звеньев различных воспринимающих (сенсорных) систем (корковых концов анализаторов).
Принято выделять в зоне того или иного анализатора проекционные, или первичные, и вторичные, поля, а также третичные поля, или ассоциативные зоны. Первичные поля получают информацию, опосредованную через наименьшее количество переключений в подкорке (в зрительном бугре, или таламусе, промежуточного мозга). На этих полях как бы спроецирована поверхность периферических рецепторов (рис. 4). В свете современных данных, проекционные зоны нельзя рассматривать как устройства, воспринимающие раздражения «точку в точку». В этих зонах происходит восприятие определенных параметров объектов, т. е. создаются (интегрируются) образы, поскольку данные участки мозга отвечают на определенные изменения объектов, на их форму, ориентацию, скорость движения и т. п.
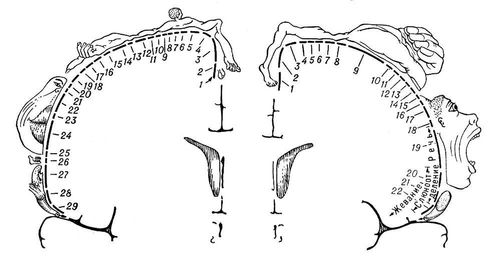
Рис. 4. Представительство чувствительных функций тела в задней центральной извилине (А) и двигательных функций — в передней центральной извилине (Б). А: 1 — половые органы; 2 — пальцы; 3 — ступня; 4 — голень; 5 — бедро; 6 — туловище; 7 — шея; 8 — голова; 9 — плечо; 10 — рука; 11 — локоть; 12 — предплечье; 13 — запястье; 14 — кисть; 15 — мизинец; 16 — безымянный палец; 17 — средний палец; 18 — указательный палец; 19 — большой палец; 20 — глаз; 21 — нос; 22 — лицо; 23 — верхняя губа; 24 — губы; 25 — нижняя губа; 26 — зубы, дёсны и челюсть; 27 — язык; 28 — глотка; 29 — внутренние органы. Б: 1 — пальцы; 2 — лодыжка; 3 — колено; 4 — бедро; 5 — туловище; 6 — плечо; 7 — локоть; 8 — запястье; 9 — кисть; 10 — мизинец; 11 — безымянный палец; 12 — средний палец; 13 — указательный палец; 14 — большой палец; 15 — шея; 16 — бровь; 17 — веко и глазное яблоко; 18 — лицо; 19 — губы; 20 — челюсть; 21 — язык; 22 — глотание. Размеры частей тела, изображенного на рисунке, соответствуют представительству двигательных и чувствительных функций организма в передней и задней центральных извилинах коры.
Кроме того, локализация функций в первичных зонах многократно дублируется по механизму, напоминающему голографию, когда каждый самый маленький участок запоминающего устройства содержит сведения о всём объекте. Поэтому достаточно сохранности небольшого участка первичного сенсорного поля, чтобы способность к восприятию почти полностью сохранилась. Вторичные поля получают проекции от органов чувств через дополнительные переключения в подкорке, что позволяет производить более сложный анализ того или иного образа. Наконец, третичные поля, или ассоциативные зоны, получают информацию от неспецифических подкорковых ядер, в которых суммируется информация от нескольких органов чувств, что позволяет анализировать и интегрировать тот или иной объект в ещё более абстрагированной и обобщённой форме. Эти области называются также зонами перекрытия анализаторов. Первичные и отчасти вторичные поля — возможный субстрат первой сигнальной системы, а третичные зоны (ассоциативные) — второй сигнальной системы, специфичной для человека (И. П. Павлов). Эти межанализаторные структуры определяют сложные формы мозговой деятельности, включающие и профессиональные навыки (нижнетеменная область), и мышление, планирование и целенаправленность действий (лобная область), и письменную и устную речь (нижняя лобная подобласть, височная, височно-теменно-затылочная и нижнетеменная области). Основные представители первичных зон в затылочной области — поле 17, где спроецирована сетчатка, в височной — поле 41, где спроецирован Кортиев орган, в прецентральной области — поле 4, где осуществляется проекция проприорецепторов в соответствии с расположением мускулатуры, в постцентральной — поля 3 и 1, где спроецированы экстерорецепторы в соответствии с их распределением в коже. Вторичные зоны представлены полями 8 и 6 (двигательный анализатор), 5 и 7 (кожный анализатор), 18 и 19 (зрительный анализатор), 22 (слуховой анализатор). Третичные зоны представлены обширными участками лобной области (поля 9, 10, 45, 44 и 46), нижнетеменной (поля 40 и 39), височно-теменно-затылочной (поле 37).
Корковые структуры играют первостепенную роль в обучении животных и человека. Однако образование некоторых простых условных рефлексов, главным образом с внутренних органов, может быть обеспечено подкорковыми механизмами. Эти рефлексы могут образовываться и на низших уровнях развития, когда ещё нет коры. Сложные условные рефлексы, лежащие в основе целостных актов поведения, требуют сохранности корковых структур и участия не только первичных зон корковых концов анализаторов, но и ассоциативных — третичных зон. Корковые структуры имеют прямое отношение и к механизмам памяти. Электрораздражение отдельных областей коры (например, височной) вызывает у людей сложные картины воспоминаний.
Характерная особенность деятельности коры — её спонтанная электрическая активность, регистрируемая в виде электроэнцефалограммы (ЭЭГ). В целом кора и её нейроны обладают ритмической активностью, которая отражает происходящие в них биохимические и биофизические процессы. Эта активность имеет разнообразную амплитуду и частоту (от 1 до 60 гц) и изменяется под влиянием различных факторов.
Ритмическая активность коры нерегулярна, однако можно по частоте потенциалов выделить несколько разных типов её (альфа-, бета-, дельтаи тета-ритмы). ЭЭГ претерпевает характерные изменения при многих физиологических и патологических состояниях (различных фазах сна, при опухолях, судорожных припадках и т. и.). Ритм, т. е. частота, и амплитуда биоэлектрических потенциалов коры задаются подкорковыми структурами, которые синхронизируют работу групп корковых нейронов, что и создаёт условия для их согласованных разрядов. Этот ритм связан с апикальными (верхушечными) дендритами пирамидных клеток. На ритмическую деятельность коры накладываются влияния, идущие от органов чувств. Так, вспышка света, щелчок или прикосновение к коже вызывают в соответствующих зонах т. н. первичный ответ, состоящий из ряда позитивных волн (отклонение электронного луча на экране осциллографа вниз) и негативной волны (отклонение луча вверх). Эти волны отражают деятельность структур данного участка коры и меняются в её различных слоях.
Филогенез и онтогенез коры. Кора — продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми структурами единый комплекс, а повреждение коры на высших этапах эволюции приводит к нарушению жизненно важных функций организма. Наибольшие изменения и увеличение в процессе эволюции новой коры претерпевают ассоциативные зоны, в то время как первичные, сенсорные поля уменьшаются по относительной величине. Разрастание новой коры приводит к вытеснению старой и древней на нижнюю и срединную поверхности мозга.
Корковая пластинка появляется в процессе внутриутробного развития человека сравнительно рано — на 2-м месяце. Раньше всего выделяются нижние слои коры (VI—VII), затем — более высоко расположенные (V, IV, III и II; см. рис. 1). К 6 месяцам у эмбриона уже имеются все цитоархитектонические поля коры, свойственные взрослому человеку. После рождения в росте коры можно выделить три переломных этапа: на 2—3-м месяце жизни, в 2,5—3 года и в 7 лет. К последнему сроку цитоархитектоника коры полностью сформирована, хотя тела нейронов продолжают увеличиваться до 18 лет. Корковые зоны анализаторов завершают своё развитие раньше, и степень их увеличения меньше, чем у вторичных и третичных зон. Отмечается большое разнообразие в сроках созревания корковых структур у разных индивидуумов, что совпадает с разнообразием сроков созревания функциональных особенностей коры. Т. о., индивидуальное (онтогенез) и историческое (филогенез) развитие коры характеризуется сходными закономерностями.
Лит.: Орбели Л. А., Вопросы высшей нервной деятельности, М.— Л., 1949; Цитоархитектоника коры большого мозга человека. Сб. ст., М., 1949; Филимонов И. Н., Сравнительная анатомия коры большого мозга млекопитающих, М., 1949; Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной деятельности животных, Полн. собр. соч., 2 изд., т. 3, кн. 1—2, М., 1951; Брейзье М., Электрическая активность нервной системы, пер. с англ., М., 1955; Сепп Е. К., История развития нервной системы позвоночных, 2 изд., М., 1959; Лурия А. Р., Высшие корковые функции человека и их нарушения при локальных поражениях мозга, М., 1962; Воронин Л. Г., Курс лекций по физиологии высшей нервной деятельности, М., 1965; Поляков Г. И., О принципах нейронной организации мозга, М., 1965; Корковая регуляция деятельности подкорковых образований головного мозга. Сб. ст., Тб., 1968; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Беритов И. С., Структура и функции коры большого мозга, М., 1969.
|
Так же Вы можете узнать о... Половые железы, гонады, органы, образующие половые клетки (яйца и сперматозоиды) у животных и человека. Процессор, центральное устройство ЦВМ, выполняющее заданные программой преобразования информации и осуществляющее управление всем вычислительным процессом и взаимодействием устройств вычислительной машины. Реннер Карл Реннер (Renner) Карл (14. |
|