Оплодотворение
Большая Советская Энциклопедия. Статьи для написания рефератов, курсовых работ, научные статьи, биографии, очерки, аннотации, описания.
|
|
| ОПА |
| ОПЕ |
| ОПЁ |
| ОПИ |
| ОПЛ |
| ОПО |
| ОПП |
| ОПР |
| ОПС |
| ОПТ |
| ОПУ |
| ОПЫ |
| ОПЬ |
Оплодотворение, сингамия, у растений, животных и человека — слияние мужской и женской половых клеток — гамет, в результате чего образуется зигота, способная развиваться в новый организм. Оплодотворение лежит в основе полового размножения и обеспечивает передачу наследственных признаков от родителей потомкам.
Оплодотворение у растений. Оплодотворение свойственно большинству растений; ему обычно предшествует образование гаметангиев — половых органов, в которых развиваются гаметы. Часто эти процессы объединяют под общим названием половой процесс. Растения, имеющие половой процесс, имеют в цикле развития и мейоз, т. е. обнаруживают смену ядерных фаз (см. Чередование поколений). Типичного полового процесса нет у бактерий и синезелёных водорослей; неизвестен он и у некоторых грибов. Типы полового процесса у низших растений разнообразны. У ряда зелёных водорослей он может осуществляться без образования гамет, в результате слияния двух одноклеточных организмов (т. н. гологамия). Слияние имеющих жгутики гамет, форма и размеры которых одинаковы, называется изогамией(см. рис. 1, 1). Этот тип полового процесса присущ многим водорослям. Одноклеточные водоросли (например, некоторые хламидомонады) как бы сами превращаются в гаметангии, образуя гаметы; у многоклеточных гаметангиями становятся некоторые клетки, не отличающиеся от других (например, у улотрикса, ульвы), или возникают морфологически отличные гаметангии (например, у эктокарпуса). Многие изогамные водоросли гетероталличны: сливаются лишь физиологически различные (+ и –) гаметы (см. Гетероталлизм). Для водорослей конъюгат (например, спирогиры) характерна конъюгация: протопласт одной клетки перетекает в другую (принадлежащую той же или др. особи), сливаясь с её протопластом (рис. 1, 2). Слияние имеющих жгутики гамет различной величины (большая — женская, меньшая — мужская; например, у некоторых хламидомонад) называется гетерогамией(рис. 1, 3). Слияние крупной безжгутиковой женские гаметы (яйцеклетка) и мелкой мужской, чаще имеющей жгутики (сперматозоид), реже — безжгутиковой (спермаций), называется оогамией. Женские гаметангии большинства оогамных низших растений называются оогониями, мужские — антеридиями. Оогамия характерна для многих зелёных, диатомовых, бурых (рис. 1, 4) и всех красных водорослей, некоторых низших грибов. У гологамных, изо-, гетерои многих оогамных растений Оплодотворение происходит в воде, у некоторых оогамных (вольвокса, вошерии) — в женских гаметангиях — оогониях, к которым вышедшие в воду сперматозоиды активно перемещаются (что, видимо, обусловлено хемотаксисом), а спермации красных водорослей — пассивно, током воды. У растений с гаметангиогамией гаметы не дифференцируются. Так, у мукоровых грибов сливаются многоядерные гаметангии, возникающие на концах выростов мицелиев (разных при гетероталлизме) (рис. 1, 5); при этом попарно сливаются и ядра. Этот тип гаметангиогамии называется зигогамией. У большинства сумчатых грибов многоядерный протопласт антеридия переливается в базальную клетку женского гаметангия (аскогон), содержащую протопласт со множеством ядер; ядра лишь попарно сближаются, образуя т. н. дикарионы (первый этап полового процесса — плазмогамия). Из аскогона вырастают гифы, в них ядра дикарионов синхронно делятся; на концах гиф возникают сумки — клетки, содержащие по дикариону. В сумках (асках) происходит второй этап полового процесса — кариогамия, т. е. слияние ядер (рис. 1, 6). Для базидиальных грибов характерна соматогамия: они не образуют ни гамет, ни гаметангиев; плазмогамия происходит у них при слиянии двух одноядерных клеток, т. н. первичных (+ и –) мицелиев; возникающая при этом двуядерная клетка даёт начало вторичному мицелию, состоящему из клеток, содержащих дикарионы; на этом мицелии образуются базидии, в них и происходит кариогамия (рис. 1, 7). Гаметангиои соматогамия — выработанное грибами в процессе эволюции приспособление к существованию вне водной среды.

Рис. 1. Оплодотворение у растений: 1 — изогамия у хламидомонады Рейнхардта; 2 — конъюгация у спирогиры; 3 — гетерогамия у хламидомонады Брауна; 4 — оогамия у фукуса: а — оогоний, б — группа антеридиев, в — сперматозоид, г — сперматозоиды близ яйцеклетки; 5 — зигогамия у мукора; 6 — дикарион (а), диплоидное ядро (б) и образование спор (в) в сумке сумчатого гриба; 7 — дикарион (а) и диплоидное ядро (б) в базидии и образование спор (в) у базидиального гриба; 8 — вскрывшийся архегоний (а), выход сперматозоидов из антеридия (б) и сперматозоиды (в) у зелёных мхов; 9 — выход сперматозоидов из антеридия (а), сперматозоид (б) и вскрывшийся архегоний (в) у равноспоровых папоротников; 10 — пыльцевое зерно (а), сперматозоид (б) и часть семезачатка саговника (в), в котором видны часть женского заростка с архегониями и мужские заростки, выдающиеся в архегониальную камеру; 11 — пыльцевое зерно (а) и часть семезачатка сосны (б), где видны часть женского заростка с архегониями и пыльцевая трубка со спермиями; 12 — пыльцевое зерно (а) и часть зародышевого мешка покрытосеменного растения (б): ядро одного спермия находится близ ядра яйцеклетки, а ядро другого — близ вторичного ядра зародышевого мешка.
Все высшие растения оогамны, но Оплодотворение у них осуществляется по-разному. Типичные гаметангии высших растений — антеридии (мужские) и архегонии (женские) многоклеточны; клетки наружного слоя гаметангия стерильны. Яйцеклетки образуются в архегониях по одной, сперматозоиды — в антеридиях, как правило, помногу. Мохои папоротникообразным для осуществления Оплодотворение необходима вода, в которой вышедшие из антеридиев сперматозоиды плывут к архегониям. Из вскрывшейся вершины готового к Оплодотворение архегония выступает слизь, привлекающая сперматозоиды. Двигаясь в слизи, сперматозоиды достигают яйцеклетки и один из них сливается с ней (рис. 1, 8 и 9). У папоротникообразных и семенных растений Оплодотворение происходит на (или в) заростке (гаметофите), существующем у первых самостоятельно, а у вторых — на спорофите. У равноспоровых папоротников заростки обоеполы, у разноспоровых и всех семенных растений раздельнополы. У семенных растений антеридиев нет: сперматозоиды (у саговников, гинкго) или безжгутиковые спермии (у всех остальных) образуются в мужских заростках (пыльцевых зёрнах). У некоторых голосеменных (гнетум, вельвичия) и всех покрытосеменных архегониев нет и яйцеклетки находятся в женских заростках. У семенных растений Оплодотворение возможно лишь после опыления — перенесения пыльцевых зёрен из микроспорангиев в пыльцевые камеры семезачатков (у голосеменных) или на рыльца пестиков (у покрытосеменных). У саговников и гинкго сперматозоиды выходят в архегониальную камеру семезачатка (рис. 1, 10) и, двигаясь в жидкости, выработанной самим растением, достигают архегониев. У семенных растений, имеющих спермии, последние перемещаются к яйцеклеткам по пыльцевым трубкам (рис. 1, 11 и 12). У покрытосеменных происходит двойное оплодотворение: один спермий сливается с яйцеклеткой, второй — с центральной клеткой зародышевого мешка (женского заростка). Осуществление Оплодотворение вне зависимости от наличия свободной воды — одно из важнейших приспособлений семенных растений к существованию на суше.
Лит.: Мейер К. И., Размножение растений, М., 1937; Навашин С. Г. Избр. труды, т. 1, М. — Л., 1951; Тахтаджян А. Л., Высшие растения, т. 1, М. — Л., 1956; Сладков А. Н., Половой процесс и жизненные циклы у растений, «Биологические науки», 1969, № 3—4.
А. Н. Сладков.
Оплодотворение у животных и человека заключается в слиянии (сингамии) двух гамет разного пола — спермия (сперматозоида) и яйца. Оплодотворение имеет двоякое значение: 1) контакт спермия с яйцом выводит последнее из заторможенного состояния и побуждает к развитию (см. Активация яйца); 2) слияние гаплоидиых ядер спермия и яйца — кариогамия — приводит к возникновению диплоидного синкариона, объединяющего отцовские и материнские наследственные факторы. Возникновение при Оплодотворение новых комбинаций этих факторов создаёт генетическое разнообразие, служащее материалом для естественного отбора и эволюции вида. Необходимая предпосылка Оплодотворение — уменьшение числа хромосом вдвое, что происходит во время мейоза. Эти деления у мужских гамет осуществляются до формирования спермия, тогда как соотношения между делениями созревания яйца и Оплодотворение у разных животных различно: спермий может проникать в яйцо до начала мейоза (губки, некоторые черви, моллюски, из млекопитающих — собака, лисица, лошадь); на стадии метафазы 1-го деления созревания (некоторые черви, моллюски, насекомые, асцидии); на стадии метафазы 2-го деления (ланцетник, многие позвоночные) и после завершения мейоза (кишечнополостные, морские ежи).
Встреча сперматозоида с яйцом обычно обеспечивается плавательными движениями мужских гамет после того, как они выметаны в воду или введены в половые пути самки (см. Осеменение). Встрече гамет способствует выработка яйцами гамонов, усиливающих движения спермиев и продлевающих период их подвижности, а также веществ, вызывающих скопление спермиев вблизи яйца. Возникновение таких скоплений у гидроидных полипов из рода Campanularia и некоторых рыб иногда рассматривают как следствие привлечения спермиев (хемотаксис), подобного наблюдаемому при Оплодотворение у мхов, папоротников и др. Однако существование направленных движений для спермиев животных недоказано; сперматозоиды двигаются беспорядочно и вступают в контакт с яйцом в результате случайного столкновения, а образование их скоплений, вероятно, вызывается действием механизма типа «ловушки», задерживающего спермии, случайно приблизившиеся к яйцу.
Зрелое яйцо окружено оболочками, имеющими у некоторых животных отверстия для проникновения спермиев — микропиле. У большинства животных микропиле отсутствует, и, чтобы достигнуть поверхности ооплазмы, спермии должны проникнуть через оболочку, что осуществляется с помощью специального органоида сперматазоида — акросомы. После того как спермий концом головки коснётся яйцевой оболочки, происходит акросомная реакция: акросома раскрывается, выделяя содержимое акросомной гранулы (рис. 2, стадии I, II), и заключённые в грануле ферменты растворяют яйцевые оболочки. В том месте где раскрылась акросома, её мембрана сливается с плазматической мембраной спермия; у основания акросомы акросомная мембрана выгибается и образует один или несколько выростов (рис. 2, стадия II) которые заполняются расположенным между акросомой и ядром (субакросомальным) материалом, удлиняются и превращаются в акросомные нити или трубочки (рис. 2, стадия III). Длина этих нитей у разных животных варьирует от 1 до 90 мкм (в зависимости от толщины барьера, который спермию приходится преодолевать). Акросомная нить проходит через растворённую зону яйцевой оболочки, вступает в контакт с плазматической мембраной яйца и сливается с ней (рис 2, стадии III, IV). У животных, спермии которых проникают в яйцо через микропиле (насекомые, головоногие моллюски, осетровые и костистые рыбы), акросома утрачивает своё первоначальное значение и иногда редуцируется или полностью исчезает (у некоторых веснянок, костистых рыб). У млекопитающих овулировавшее яйцо, кроме оболочки, окружено несколькими слоями фолликулярных клеток яйценосного бугорка. У лошади, коровы, овцы фолликулярные клетки рассеиваются вскоре после овуляции, и спермий свободно достигает поверхности яйцевой оболочки. У большинства млекопитающих клетки яйценосного бугорка сохраняются на протяжении нескольких часов и, чтобы проникнуть через этот барьер, спермии выделяют фермент гиалуронидазу, который растворяет вещество, связывающее фолликулярные клетки между собой. Гиалуронидаза, как и фермент, растворяющий яйцевую оболочку, заключена в акросоме. Сразу после эякуляции спермии неспособны к выделению этих ферментов; такая способность возникает под действием содержимого женских половых путей, вызывающего определённые физиологические изменения спермиев (процесс капацитации).
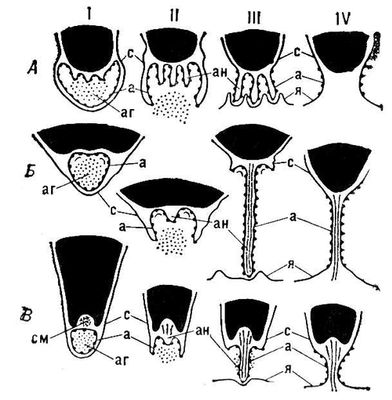
Рис. 2. Схема осуществления акросомной реакции: А — у кольчатого червя; Б — у кишечнодышащего; В — у морского ежа; I—IV — последовательные стадии реакции; а — акросомная мембрана, аг — акросомная гранула, ан — акросомная нить, с — плазматическая мембрана спермия, см — субакросомальный материал, я — плазматическая мембрана яйца.
С момента слияния плазматических мембран гамет в месте контакта акросомной нити с поверхностью ооплазмы яйцо и спермий — уже единая клетка — зигота. Вскоре обнаруживаются первые признаки активации яйца: кортикальная реакция и стягивание ооплазмы в месте контакта с акросомной нитью спермия, приводящее к образованию воспринимающего бугорка. Ооплазма этого бугорка обтекает ядро, центриоли и митохондрии сперматозоида, а иногда и осевой стержень его хвоста, вовлекая их в глубь яйца, тогда как плазматическая мембрана спермия остаётся на поверхности и встраивается в плазматическую мембрану яйца, так что поверхностная мембрана зиготы имеет мозаичное строение. Погрузившись в ооплазму, головка спермия поворачивается на 180°, и у её основания формируется сперматическая звезда (рис. 3, стадии I, II). Постепенно головка набухает и преобразуется в пузыревидный мужской пронуклеус, перемещающийся вслед за сперматической звездой, которая как бы увлекает его за собой (рис. 3, стадии II, III, IV). Мужской пронуклеус сближается с женским, а сперматическая звезда делится на две, участвующие затем в образовании веретена 1-го деления дробления. У кишечнополостных, плоских червей, морских ежей пронуклеусы сливаются в единое ядро зиготы (рис. 3, стадия V), у некоторых круглых червей, моллюсков, ракообразных, рыб и земноводных они длительное время остаются в тесном контакте, но не сливаются, и объединение отцовского и материнского ядерного материала происходит только на стадии метафазы 1-го деления дробления. Одновременно с этими изменениями в яйце повышается интенсивность обмена веществ: увеличивается проницаемость клеточной мембраны, активируется синтез белка и др.

Рис. 3. Оплодотворение у морского ежа: I—IV — последовательные стадии преобразования спермия и формирования мужского пронуклеуса; V — слияние пронуклеусов; з — сперматическая звезда, м — митохондрия из средней части спермия, п — женский пронуклеус, пт — полярные тельца, х — хвостовой отдел спермия, я — ядро спермия.
При Оплодотворение яиц животных с наружным осеменением в яйцо проникает только один спермий (физиологическая моноспермия); это обеспечивается особым механизмом, в основе которого лежит процесс секреции содержимого кортикальных телец, предотвращающий проникновение спермиев в ооплазму. Среди животных с внутренним осеменением наряду с моноспермными встречаются и такие, у которых в яйцо проникает несколько спермиев (физиологическая полиспермия); однако и в этих случаях с женским пронуклеусом сливается только одно сперматическое ядро. См. также Наследственность и Пол.
Лит.: Ротшильд Н. М., Оплодотворение, пер. с англ., М., 1958; Дорфман В. А., Физико-химические основы оплодотворения, М., 1963; Гинзбург А. С., Оплодотворение у рыб и проблема полиспермии, М., 1968; Austin С. R., Fertilization, Englewood Cliffs (N. J.), 1965; Fertilization, ed. Ch. B. Metz, A. Monroy, v. 1—2, N. Y. — L., 1967—69; Reproduction in mammals, ed. C. R. Austin, R. V. Short, book 1 — Germ cells and fertilization, L., 1972.
А. С. Гинзбург.
|
Так же Вы можете узнать о... Исламбеков Раджаб Капланович (р. 30.12.1925, Ташкент), советский эндокринолог, член-корреспондент АМН СССР (1969), заслуженный деятель науки Узбекской ССР (1964). Карпентер Малколм Скотт Карпентер (Carpenter) Малколм Скотт (р. 1.5. Кнудсен Мартин Ханс Кристиан Кнудсен (Knudsen) Мартин Ханс Кристиан (15.2. Коротнёв Алексей Алексеевич [15(27).2.1854, Москва, — 14(27). Курдючные овцы, породы грубошёрстных и полугрубощёрстных овец мясо-сального типа, имеющих жировые отложения на крестце (курдюк). Лимонит (нем. Limonit, от греч. leimon — луг; сырое место), собирательное название для природных скрытокристаллических, частью аморфных минеральных агрегатов, представляющих собой смеси минералов — гидроокислов трёхвалентного железа (гётита, гидрогётита, гидрогематита, лепидокрокита). Малипьеро Джан Франческо Малипьеро (Malipieго) Джан Франческо (родился 18. Металлогения (от металлы и греч. –géneia — часть сложного слова, означающая происхождение, создание), раздел учения о полезных ископаемых, исследующий региональные закономерности формирования и размещения рудных месторождений. Московский электромеханический завод им. Владимира Ильича, одно из старейших машиностроительных предприятий СССР. Нереис (Nereis), род многощетинковых червей семейства нереид. Окорка древесины, очистка от коры необработанных лесоматериалов. Папеэте (Papeete), город, административный центр французских владений в Океании (Французская Полинезия, официально заморская территория), на острове Таити (острове Общества). Пищевой промышленности институты, втузы, готовящие инженерные и научные кадры для пищевой промышленности. Правёж, по древнерусскому феодальному праву способ исполнения судебного решения, принуждение к уплате долгов, пошлин и др. |
|